فصل چهارم: سیناپس و انتقال پیامها بین نورونها
محل ارتباط یک نورون را با نورون دیگر سیناپس میخوانند. این ارتباط به وسیله غشای نورونها صورت میگیرد. بیشترین تعداد سیناپسها از نوع آکسون به دندریت و یا آکسون به جسم سلولی است؛ ولی در برخی نواحی مغز و نخاع حالتهای دیگری مانند سیناپسهای آکسون به آکسون و دندریت به دندریت نیز شناسایی شده است. در هر حال ساختمان و طرز عمل کلی سیناپسها در همه حالات شبیه به هم است. انتهای آکسون به شاخه های زیادی تقسیم میشود و هر شاخه به بخشی به نام گره یا دکمه سیناپس ختم میشود که کمی برجسته تر است. این ناحیه با غشای نورون بعدی سیناپس می سازد. نورون اول را پیش سیناپسی و نورون بعدی را پس سیناپسی می خوانند. در دستگاه عصبی مرکزی هر نورون ممکن است نسبت به برخی نورونها پیش سیناپسی و نسبت به برخی دیگر پس سیناپسی باشد.
سیناپسهای الکتریکی و سیناپسهای شیمیایی
مجموعه سیناپسهایی که تاکنون در دستگاه عصبی جانوران شناسایی شده اند به دو گروه الکتریکی و شیمیایی تقسیم میشوند. در سیناپسهای الکتریکی که در دستگاه عصبی بی مهرگان یافت میشود غشای دو نورون پیش سیناپسی و پس سیناپسی به یکدیگر می چسبد و فاصله ای بین آنها وجود ندارد. موج عصبی پس از رسیدن به این نوع سیناپس بدون واسطه به غشای بعدی انتقال می یابد و این انتقال به واسطه شیمیایی نیاز ندارد و اغلب در هر دو جهت امکان پذیر است. ضمناً سیناپسهای الکتریکی همواره از نوع تحریک کننده است. زمان عبور جریان از سیناپسهای الکتریکی با توجه به فقدان انتقال دهنده شیمیایی در آنها بسیار کوتاه است. اکثر سیناپسها در دستگاه عصبی مهره داران و بی مهرگان از نوع شیمیایی است. بررسی با میکروسکوپ الکترونی نشان داده است که در این سیناپسها غشاهای دو نورون پیش سینا پسی و پس سیناپسی به یکدیگر نمی چسبند و در بین آنها فاصله ای در حدود ۲۰۰ تا ۳۰۰ آنگستروم وجود دارد. این ناحیه را فضا یا شکاف سیناپسی می نامند. ساختمان غشا در محل سیناپس با سایر نواحی غشای نورون متفاوت است. در درون دکمه سیناپسی تعدادی کیسه های ترشحی بسیار ریز وجود دارد که در آنها واسطه شیمیایی سیناپس یافت میشود؛ علاوه بر آن برخی ضمایم سیتوپلاسم به خصوص تعدادی میتوکندری نیز در این ناحیه وجود دارد میتوکندریها انرژی لازم را برای کار دکمه سیناپسی فراهم میکنند. غشای نورونها در بعضی سیناپسها صاف و در برخی دیگر بسیار چین خورده است و بر روی آنها گیرنده های غشایی خاصی وجود دارد که در تنظیم آزاد شدن میانجی شیمیایی و اثر آن در نورون پس سیناپسی نقش دارند. میکروسکوپ الکترونی وجود رشته ها و لوله های بسیار باریک را در شکاف سیناپس نشان داده است. این رشته ها و لوله ها غشاهای دو نورون را به یکدیگر مربوط و به هدایت انتقال دهنده به سوی گیرنده های آن کمک می کنند. انتقال موج عصبی در سیناپس شیمیایی در حدود ۰/۵ تا یک هزارم ثانیه طول می کشد.
سیناپسهای تحریک کننده و سیناپسهای بازدارنده
سیناپسهای شیمیایی به دو نوع تحریک کننده و بازدارنده تقسیم می شوند. امکان بازداری و مهار نورون پس سیناپسی یکی از مزایای سیناپس شیمیایی است و در سیناپسهای الکتریکی وجود ندارد. رسیدن موج عصبی به سیناپس و آزاد شدن انتقال دهنده عصبی در شکاف سیناپس ممکن است باعث انتقال موج عصبی به نورون بعدی شود که در این صورت سیناپس از نوع تحریک کننده است. در برخی از سیناپسهای دیگر رسیدن موج عصبی و آزاد شدن انتقال دهنده در شکاف سیناپس نه تنها نورون بعدی را تحریک نمیکند بلکه آستانه تحریک آن را بالا می برد و احتمال تحریک آن را از طریق سایر سیناپسها نیز کاهش میدهد. چنین سیناپسهایی را بازدارنده یا مهار کننده میخوانند. تحریکی یا مهاری بودن یک سیناپس با نوع انتقال دهنده و نوع گیرنده آن بستگی دارد؛ به همین جهت یک انتقال دهنده ممکن است در بعضی سیناپسها نقش تحریکی و در برخی دیگر نقش مهاری داشته باشد.
هنگامی که موج عصبی به دکمه سیناپسی میرسد و غشای پیش سیناپسی را دپولاریزه می کند تغییراتی در ساختمان غشای آن ناحیه رخ میدهد که طی آن کلسیم به پایانه سیناپس وارد میشود و انتقال دهنده از دکمه سیناپسی به شکاف سیناپس آزاد می گردد. روند آزاد شدن انتقال دهنده از دکمه سیناپسی به صورت کوانتایی صورت می گیرد؛ یعنی به ازای هر پتانسیل عمل که به پایانه سیناپس می رسد مقدار معین و ثابت انتقال دهنده به فضای سیناپس رها میشود؛ بنابراین هر چه تعداد پتانسیلهای عملی که به سیناپس می رسد بیشتر باشد مقدار انتقال دهنده آزاد شده نیز زیادتر خواهد شد. به همین دلیل است که تحریک مکرر و شدید نورونها میتواند باعث خستگی سیناپس شود. در این حالت نورون فرصت بازسازی انتقال دهنده های عصبی را پیدا نمی کند و سیناپس بر اثر کمبود این مواد از کار می افتد. در شرایط طبیعی و مناسب تحریک مکرر و شدید تار عصبی به شرط رعایت زمان مرحله تحریک ناپذیری آن باعث خستگی تار عصبی نمیشود؛ در حالی که انتقال پیامها در سیناپسها متوقف می شود. انتقال دهنده های عصبی پس از آزاد شدن و اثر در غشای پس سیناپسی به وسیله آنزیمهایی تجزیه یا دوباره جذب پایانه های سیناپسی میشوند.
طرز کار سیناپسهای تحریک کننده
همان طور که قبلاً اشاره شد رسیدن موج عصبی پتانسیل عمل به انتهای آکسون و دپولاریزه شدن غشای دکمه سیناپسی باعث تراوا شدن آن به یونهای کلسیم و ورود تعدادی یون کلسیم به درون پایانه سیناپس میشود. ورود کلسیم باعث اتصال غشای پیش سیناپسی به غشای تعدادی از حفره های محتوی انتقال دهنده و سرانجام آزاد شدن محتویات این حفره ها به درون شکاف سیناپس میشود. در سیناپسهای تحریک کننده، اثر انتقال دهنده در گیرنده های غشای پس سیناپسی باعث نفوذ پذیر شدن این غشا نسبت به یونهای سدیم می شود. باز شدن مجاری سدیم در این غشا و ورود تعدادی یون سدیم باعث کاهش پتانسیل غشا و در نتیجه تحریک آن می گردد.
طرز کار سیناپسهای بازدارنده
در این سیناپسها آزاد شدن انتقال دهنده عصبی و اثر آن در گیرنده های غشای پس سیناپسی تأثیری در نفوذ پذیری آن نسبت به یون سدیم ندارد؛ ولی نفوذ پذیری غشای مذکور را نسبت به یونهای پتاسیم و یا کلر افزایش می دهد که هر یک از دو حالت مذکور به افزایش پتانسیل آرامش غشای پس سیناپسی منجر میشود. اگر اثر انتقال دهنده در غشای پس سیناپسی، نفوذ پذیری آن را نسبت به پتاسیم افزایش دهد با توجه به تراکم بیشتر این یون در داخل نورون نسبت به خارج مقدار بیشتر پتاسیم به بیرون انتشار می یابد و بر بارهای مثبت خارج افزوده از بارهای مثبت درون میکاهد. در صورتی که اثر انتقال دهنده در غشای پس سیناپسی باعث افزایش تراوایی آن نسبت به یون کلر شود با توجه به تراکم بیشتر کلر در مایع خارج نورون، تعدادی یون کلر به درون انتشار یافته بر بارهای منفی داخل می افزاید و از بارهای منفی خارج میکاهد. درباره سیناپسهای بازدارنده باید به نقش مهار پیش سینا پسی نیز توجه داشت. این پدیده در برخی سیناپسهای مغز و نخاع باعث توقف عبور موج عصبی در سطح پیش سیناپسی میشود و به واسطه سیناپسهای آکسون به آکسون صورت میگیرد. در این حالت دکمه سیناپسی یک نورون رابط در مجاورت دکمه سیناپسی یک نورون تحریک کننده قرار می گیرد و انتقال دهنده ای آزاد میکند که نفوذ پذیری غشای نورون تحریک کننده را نسبت به یون کلسیم کاهش میدهد. در این صورت با رسیدن موج عصبی به انتهای نورون تحریک کننده کلسیم کافی وارد پایانه نشده و خروج انتقال دهنده کاهش می یابد و در نتیجه موج عصبی در سیناپس متوقف میشود.
گیرنده های پیش سینا پسی و پس سیناپسی
هر یک از دو غشای پیش سیناپسی و پس سیناپسی دارای گیرنده هایی خاص است. گیرنده های غشایی از نوع ترکیبات پروتئینی است. بخشی از آن که به طرف خارج در درون فضای سیناپس قرار دارد با انتقال دهنده ترکیب میشود و بخش دیگر گیرنده در عرض غشا و درون نورون قرار گرفته است و در تغییر نفوذ پذیری غشا و تغییر سوخت و ساز نورون شرکت میکند. برخی از گیرنده های پیش سیناپسی نسبت به انتقال دهنده ای که از همان پایانه عصبی آزاد میشود حساسیت دارند. این گیرنده ها با یک چرخه بازخورد منفی باعث تنظیم مقدار انتقال دهنده عصبی میشوند و مانع آزاد شدن بیش از حد آن میشوند. برخی دیگر گیرنده هایی هستند که نسبت به سایر انتقال دهنده ها یا مواد شیمیایی حساسیت دارند. گیرنده های پس سیناپسی در تحریک و یا مهار پس سیناپسی نقش دارند و اثر انتقال دهنده در آنها نفوذ پذیری غشای پس سینا پسی را به سدیم یا پتاسیم یا کلر افزایش میدهد و یا باعث تغییر متابولیسم نورون پس سیناپسی میشود. دکمه سیناپسی مانند یک مبدل انرژی عمل می کند و در آن با واسطه مواد شیمیایی انرژی الکتریکی پتانسیل عمل یک نورون در نورون بعدی به تغییرات بیوالکتریک منجر میشود. اثر انتقال دهنده در گیرنده پس سیناپسی خود اغلب مانند قرار گرفتن یک کلید در قفل است که باعث باز شدن مجرای عبور یون یا یونهای خاصی میشود.
برخی از گیرنده های غشایی به طور مستقیم باعث باز شدن مجاری یونها نمیشوند بلکه اثر انتقال دهنده در آنها باعث فعال شدن نوکلئوتیدهای حلقوی می شود. در این حالت ماده ای به نام آدنوزین مونوفسفات حلقوی به وجود می آید و به وسیله آنزیمهایی به نام پروتئین کیناز شکل پروتئینهای مجاری غشای پس سیناپسی را تغییر میدهند و باعث عبور یونها و تغییر موضعی پتانسیل غشا میشوند. با توجه به آنچه گفته شد گیرنده های نوروترانسمیترها را می توان به دو گروه یونوتروپیک و متابو تروپیک تقسیم کرد. گیرنده های یونوتروپیک از نوع کانالهای یونی دریچه دار وابسته به لیگاند هستند یعنی باز شدن آنها با تغییرات ولتاژ غشا صورت نمی گیرد و مربوط به اثر یک ماده خاص بر غشا است. اثر نوروترانسمیتر بر این گیرنده ها باعث تغییر شکل فضایی آنها و باز شدن مجرای میان گیرنده شده غشای نورون را برای عبور یونها تراوا میکند. در گیرنده های متابو تروپیک، اثر نوروترانسمیتر موجب راه اندازی یک سری تغییرات شیمیایی در نورون پس سیناپسی میشود که در نهایت به تغییر پتانسیل غشای پس سیناپسی منجر میشود. در این مسیر که پیچیده تر و کندتر است تغییرات ایجاد شده دیرتر پدیدار شده و اثر آنها طولانی تر است.
هم گرایی و واگرایی سیناپسی
هر یک از نورونهای مغز یا نخاع با تعداد زیادی نورون دیگر سیناپس دارد. هر نورون در مراکز عصبی ممکن است نسبت به تعدادی نورون پیش سیناپسی و نسبت به عده ای دیگر پس سیناپسی باشد. از مجموع پایانه های سیناپسی که در یک نورون اثر می گذارند برخی تحریک کننده و برخی دیگر بازدارنده اند. پایانه های تحریک کننده با اثر خود پتانسیل آرامش غشای پس سیناپسی را کاهش و پایانه های بازدارنده با اثر خود پتانسیل آرامش غشای پس سیناپسی را افزایش میدهند. این دو اثر به ترتیب پتانسیل پس سیناپسی تحریک کننده (EPSP) و پتانسیل سیناپسی بازدارنده (IPSP) خوانده میشوند. نتیجه دو پدیده مذکور باعث می شود تا نورون پس سیناپسی به کار افتد و یا از کار بازماند.
انتقال دهنده های عصبی
نورونها از نظر تولید و ترکیب مواد شیمیایی یاخته هایی بسیار فعال اند و فراورده های آنها در سوخت و ساز آنها ایجاد ارتباط بین نورونها کار کردهای مختلف عصبی مانند حالات روانی و یادگیری نقش اساسی دارند. بعضی از این مواد در سیناپسها به عنوان انتقال دهنده و برخی به عنوان تعدیل کننده عمل میکنند. بسیاری از داروهای روانی با تأثیر در این مواد و یا در گیرنده های آنها باعث تغییرات عصبی و روانی می گردند. انتقال دهنده که در سطح یک سیناپس اثر میکند منطقه عمل محدودتری دارد. تعدیل کننده های عصبی در مغز میدان عمل وسیع تری دارند و میزان ترشح آنها بیشتر است. ساخته و آزاد شدن این مواد به تولید و ترشح هورمونها شباهت دارد. تاکنون دهها انتقال دهنده و تعدیل کننده عصبی شناسایی شده اند و احتمالاً تعداد این مواد بسیار بیش از آن است که تا به حال شناخته شده اند. انتقال دهنده ها و تعدیل کننده ها را میتوان به دو دسته مواد کوچک مولکول و بزرگ مولکول تقسیم کرد. میانجیهای کوچک مولکول شامل اسیدهای امینه از قبیل گابا گلیسین و گلوتامات استیل کولین و مونو آمینها از قبیل اپی نفرین نوراپی نفرین دوپامین و سروتونین و آدنوزین تری فسفات (ATP)) است. میانجیهای بزرگ مولکول از نورو پپتیدها مانند ماده ، آنکفالین و واز و پرسین تشکیل شده اند. در اینجا به ذکر مهم ترین انتقال دهنده ها و تعدیل کننده های عصبی می پردازیم.
۱) استیل کولین
این ماده که از ترکیب کولین با استیل کوآنزیم A به وجود می آید اولین انتقال دهنده عصبی است که در اوایل قرن بیستم کشف شد. استیل کولین انتقال دهنده منحصر به فرد همه سیناپسهای عصب به عضلات اسکلتی است. علاوه بر آن در عقده های خودکار و پایانه های سیناپسی اعصاب پاراسمپاتیک و بخشی از اعصاب سمپاتیک نیز این ماده آزاد میشود. آزاد شدن استیل کولین مانند سایر انتقال دهنده ها با دپولاریزه شدن دکمه سیناپسی و سپس ورود کلسیم به آن صورت می گیرد. نورونها و سیناپسهایی که استیل کولین آزاد میکنند استیل کولینرژیک خوانده می شوند. استیل کولین در تارهای عضلات اسکلتی اثر تحریکی و در تارهای عضلاتی قلب اثر بازدارندگی دارد و این تفاوت به نوع گیرنده های پس سیناپسی مربوط است؛ به این ترتیب که استیل کولین در عضلات اسکلتی در مجاری سدیم اثر کرده باعث دپولاریزاسیون تارهای عضلانی میشود در حالی که گیرنده های استیل کولین در قلب مربوط به مجاری یونهای پتاسیم بوده اثر استیل کولین در آنها باعث افزایش پتانسیل آرامش و کاهش تحریک پذیری آنها میشود.
استیل کولین دارای دو نوع گیرنده موسکارینی و نیکوتینی است. گیرنده های موسکارینی به وسیله استیل کولین و ماده ای به نام موسکارین که از یک قارچ سمی به دست میآید فعال میشوند. گیرنده های نیکوتینی را استیل کولین و نيكوتين، فعال میسازند. آتروپین و کورار به ترتیب گیرنده های موسکارین و نیکوتین را از کار می اندازند. در سیناپسهای استیل کولینرژیک مغز و نخاع از هر دو نوع گیرنده های موسکارینی و نیکوتینی یافت میشود؛ ولی گیرنده های تارهای عضلات اسکلتی و عقده های گانگلیون) نباتی اتونوم) با دو زیرگروه از نوع نیکوتینی هستند. نقش استیل کولین در مغز در بسیاری از اعمال عصبی مانند حافظه یادگیری و تنظیم مراحل خواب نشان داده شده است. در بیماری آلزایمر که با زوال عقل، اختلال در حافظه و یادگیری همراه است نورونهای استیل کولینرژیک مغز دچار کم کاری و مرگ تدریجی میشوند. گیرنده های موسکارینی استیل کولین دارای پنج زیر گروه اند و در نواحی مختلف دستگاه عصبی مرکزی، قلب، غدد و عضلات صاف قرار دارند. اثر استیل کولین بر گیرنده های نیکوتینی (عضلات اسکلتی، نورونهای پیش عقده ای سمپاتیک پاراسمپاتیک و برخی سیناپسهای دستگاه عصبی مرکزی همیشه تحریکی است ولی اثر استیل کولین بر گیرنده های موسکارینی ماهیچه صاف و قلبی غدد درون ریز و برون ریز و برخی سیناپسهای دستگاه عصبی مرکزی تحریکی یا مهاری است. رسپتورهای نیکوتینی از نوع یونوتروپیک و رسپتورهای موسکارینی از نوع متابو تروپیک است.
استیل کولین پس از آزاد شدن از دکمه سیناپسی به وسیله آنزیم استیل کولین استراز تجزیه و غیر فعال و بخشی از مواد اولیه آن دوباره جذب پایانه سیناپس میشود. اثر بسیاری از داروها و مواد شیمیایی مؤثر در اعصاب با تأثیر آنها در ترشح استیل کولین یا اثر استیل کولین در گیرنده های آن مربوط است. داروهای کولینرژیک باعث تسهيل عمل استیل کولین و داروهای آنتی کولینرژیک مانع عمل آن می شوند. یک ماده سمی به نام توکسین بوتولیک که از نوعی باکتری تولید می شود مانع آزاد شدن استیل کولین و در نتیجه از کار افتادن سیناپسهای کولینرژیک میشود. آفت کشها و ترکیبات ارگانوفسفات و گازهای عصبی که در جنگهای شیمیایی به کار میروند به سرعت از پوست ریه ها و ملتحمه چشم جذب شده و با خنثی کردن اثر استیل کولین استراز باعث طولانی شدن فعالیت سیناپسهای کولینرژیک میشود که به تشنج و اغما منجر میگردد. آتروپین به همراه کاربامات و پیریدوستیگمین به عنوان پادزهر مواد آنتی کولین استرازیک در سربازان و سایر افرادی که ممکن است در معرض گازهای عصبی قرار گیرند به کار میرود.
۲) کاتکولامینها
این مواد شامل دوپامین نوراپی نفرین (نورآدرنالین) و اپی نفرین (آدرنالین) هستند که همگی از اسید آمینه تیروزین ساخته می شوند.
الف) دوپامین
این ماده یکی از مهمترین انتقال دهنده های نورونهای مراکز عصبی است و بسیاری از بیماریهای عصبی و روانی به اختلال در ترشح و عملکرد آن مربوط است. دوپامین بر حسب نوع گیرنده هایی که در آن اثر می کند ممکن است تحریک کننده یا بازدارنده باشد. در نواحی مختلف مغز نورونهای دوپامینرژیک شناسایی شده اند که یکی از مهمترین آنها هسته سیاه در مغز میانی است. دوپامین از انتهای آکسونهای نورونهایی که جسم سلولی آنها در هسته سیاه قرار دارد به درون اجسام مخطط در هسته های قاعده ای مغز آزاد می شود (مسیر نیگر و استریاتال. آسیب دیدن این نورونهای دوپامینرژیک باعث بیماری پارکینسون میشود. در این بیماری تحلیل تدریجی نورونهای دوپامینرژیک هسته سیاه باعث کاهش دوپامین در هسته های قاعده ای مغز میشود که این امر خود سبب پرکاری نورونهای استیل کولینرژیک در این عقده ها میگردد و لرزش و سختی عضلات و ناتوانی در حفظ تعادل و انجام حرکات طبیعی پدید می آید.
یک مسیر دیگر دوپامینرژیک مغز از ناحیه تكمنتال شکمی واقع در مغز میانی به هسته آکومبنس در هسته های قاعده ای و نواحی زیر قشری دستگاه کناری سیستم لیمبیک) مانند بادامه می رود و در رفتارهای پاداشی و اعتیاد و اختلالات روانی نقش دارد. در بیماری روان گسیختگی (اسکیزوفرنی نیز نورونهای دوپامینرژیک مغز در گیرند و به نظر می رسد که فعالیت بیش از حد طبیعی این نورونها و یا حساسیت زیاد گیرنده های آنها عامل بخشی از نشانه های اسکیزوفرنی باشد. اسکیزوفرنی با روان پریشی، توهم هذیان و اختلال در تفکر منطقی همراه است. شواهد زیادی نشان میدهند که اختلال در سیستم دوپامینرژیک مزوکورتیکال مغز میانی به قشر مخ باعث ایجاد دست کم بخشی از نشانه های اسکیزوفرنی میشود. استفاده از آمفتامینها که باعث تحریک بیش از حد بعضی گیرنده های دوپامین در سیستم لیمبیک می شود عوارض مشابه اسکیزوفرنی ایجاد میکند. احتمالاً در این بیماری تعداد گیرنده های مذکور و یا حساسیت آنها در مغز افزایش مییابد. آن دسته از داروهایی که باعث افزایش دوپامین در مغز میشوند نشانه های بیماری پارکینسون را کاهش میدهند و داروهای نورولپتیک که گیرنده های دوپامین را بازداری کرده و موجب کاهش فعالیت نورونهای دوپامینرژیک میگردند نشانه های اسکیزوفرنی را تخفیف میدهند.
معمولاً بیماریهای پارکینسون و اسکیزوفرنی را دو قطب کاهش و افزایش فعالیت دوپامین در مغز میدانند. اثر اغلب داروهای ضد روان پریشی پیشگیری از تحریک شدن گیرنده های دوپامین است. از حدود ۱۹۵۰ به بعد به تدریج داروهایی مانند کلرپرومازین هالوپریدول پرفنازین و فلوفنازین برای درمان اسکیزوفرنی ساخته شده اند و در اواخر قرن بیستم داروهای جدیدتری مانند کلوزاپین رسپیریدون و اولانزاپین بر آنها افزوده شده است که همگی به رفع اختلالات انتقال دهنده های درگیر در روان پریشی کمک میکنند. برای دوپامین رسپتورهای متعددی شناسایی شده است که هر کدام در اعمال خاصی دخالت دارند دوپامین و سایر کاتکولامینها به وسیله یک آنزیم داخل نورون به نام مونو آمین اکسیداز (MAO) و یک آنزیم خارج نورون به نام کاتکول اومتیل ترانسفراز (COMT) تجزیه و بدون اثر می شوند. دوپامین پس از تجزیه شدن ماده ای به نام هو مو وانیلیک اسید (HVA) به وجود می آورد که با اندازه گیری مقدار آن در مایع مغزی نخاعی، پلاسمای خون و ادرار می توان به میزان فعالیت نورونهای دوپامینرژیک مغز پی برد. دوپامین در نظام پاداش مغز و احساس نشاط و سرخوشی نقش دارد و برخی از مواد مخدر اعتیاد آور مانند کو کائین باعث افزایش آزاد شدن این انتقال دهنده میشود. تاکنون پنج نوع گیرنده دوپامینی شناسایی شده است.
(ب) نوراپی نفرین (نورآدرنالین)
آنزیمی به نام دوپامین هیدروکسیلاز باعث تبدیل دوپامین به نوراپی نفرین میشود. نورونهای دوپامینرژیک فاقد این آنزیم اند؛ ولی در نورونهایی که این آنزیم را دارند دوپامین تبدیل به نوراپی نفرین و آزاد می شود. چنین نورونهایی را نور آدرنرژیک میخوانند. نوراپی نفرین در اعصاب محیطی سمپاتیک و تعدادی از سیناپسهای مغز نقش انتقال دهنده دارد. نوراپی نفرین پس از اثر در غشای پس سیناپسی دوباره به وسیله پایانه سیناپس جذب میشود و یا به وسیله آنزیم مونو آمین اکسید از تجزیه و غیر فعال میگردد. بیشتر نورونهای نورآدرنرژیک در تنه مغزی از جمله هسته نیلی لوکوس سرولئوس) در پل مغز قرار دارند. این نورونها در اغلب نواحی نقش تحریک کننده و در برخی سیناپسهای دیگر اثر بازدارنده دارند. نورونهای نورآدرنرژیک مغز در بیداری و افزایش سطح برانگیختگی و بالا بردن فعالیت نورونهای مغز شرکت دارند. گیرنده های نوراپی نفرین به دو گروه کلی آلفا و بتا که هر کدام زیر گروههایی نیز دارند تقسیم می شوند. در اغلب موارد اثر نوراپی نفرین در گیرنده های آلفا از نوع تحریکی و در گیرنده های بتا از نوع مهاری است. قلب و روده وضعی استثنایی دارند. نور آدرنالین در گیرنده های بتا در قلب اثر تحریک کننده دارد و اثر نورآدرنالین در هر دو نوع گیرنده های آلفا و بتا در روده باعث مهار حرکات روده می شود.
(ج) اپی نفرین (آدرنالین)
این ماده بیشتر در بخش مرکزی غده فوق کلیه ترشح می شود. بخش مذکور از نظر جنینی یک گره سمپاتیک تغییر شکل یافته و هورمونهای آن نیز همان موادی است که اعصاب سمپاتیک از انتهای خود آزاد می کنند. اپی نفرین از نوراپی نفرین ساخته میشود و آن دسته از نورونها که این انتقال دهنده را می سازند آدرنرژیک خوانده میشوند. اپی نفرین و سایر كاتكولامینها پس از آزاد شدن در شکاف سیناپس به میزان زیاد دوباره جذب پایانه سیناپس میشوند و مجدداً به کار میروند و فقط مقدار کمی از آنها به وسیله آنزیمهای تجزیه کننده به مواد شیمیایی غیر فعال تجزیه می شوند.
سروتونین
این ماده از اسید آمینه تریپتوفان ساخته می شود. سروتونین و ماده دیگری به نام ملاتونین با نام کلی ایندولامین خوانده می شوند. ملاتونین هورمون غده کاجی (پینه آل) است و از تغییر شکل سروتونین به وجود می آید. سروتونین که نام شیمیایی آن ۵ - هیدروکسی تریپتامین است یکی از مهم ترین انتقال دهنده ها و تعدیل کننده های عصبی است که علاوه بر غده کاجی در هسته سجافی (رافه) تنه مغزی مخچه هیپوتالاموس و نواحی دیگر دستگاه عصبی ساخته می شود. آنزیم مونو آمین اکسید از علاوه بر تجزیه کاتكولامينها باعث تجزيه و بی اثر شدن سروتونین نیز میشود. بر اثر تجزیه سروتونین ماده ای به نام ۵ - هیدروکسی ایندول استیک اسید به وجود می آید که با اندازه گیری مقدار آن در ادرار میتوان به میزان سروتونین بدن پی برد. سروتونین در مسیرهای انتقال درد به مراکز عصبی به صورت یک ماده مهار کننده عمل میکند. با توجه به افزایش فعالیت نورونهای سروتونینرژیک هسته رافه در هنگام خواب سروتونین را در برقراری حالت خواب مؤثر میدانند. سروتونین علاوه بر درد و خواب در تنظیم رفتارهای تغذیه و نوسانات خلق و خو نیز نقش دارد. اثر درمانی برخی مواد دارویی ضد افسردگی به اثر آنها در سیناپسهای سروتونینرژیک مغز مربوط است. بازدارنده های مونو آمین اکسیداز که از داروهای ضد افسردگی هستند باعث کاهش تجزیه سروتونین در سیناپسهای مغز میشوند. بعضی دیگر از مواد ضد افسردگی مانند فلوئوکستین که با نام تجارتی پروزاک شهرت یافته است به صورت اختصاصی بازجذب سروتونین را کاهش میدهد و باعث افزایش مقدار آن در سیناپسها می شود. برخی از مواد هذیان آور و توهم زا مانند ال اس دی دی اتیل امید اسید لیزرژیک که در کفک انگل خوشه های چاودار یافت می شود سیناپسهای سروتونین زا را به طور موقت غیر فعال میکنند. داروهایی که نورونهای سروتونینرژیک را فعال میکنند دوره های خواب دیدن را کاهش میدهند. پسیلوسیبین که در برخی قارچها یافت میشود و مسکالین که از گونه های کاکتوس به دست می آید و از مواد توهم زا میباشند و اکستازی یا مت آنکفالین که به طور موقت باعث سرخوشی شده و به دنبال آن موجب اختلال در تمرکز و افسردگی میگردد از موادی هستند که در رهاسازی سروتونین نیز اثر می کنند. سرخوشی ناشی از سوء مصرف اکستازی احتمالاً مربوط به رهاسازی سروتونین زیاد در سیناپسها است که متعاقب آن کمبود سروتونین موجب افسردگی میشود.
گابا (گاما آمینو بوتیریک اسید)
این ماده از اسید گلوتامیک ساخته می شود و مهم ترین واسطه شیمیایی بازدارنده اعصاب است که هم در مهار پیش سیناپسی و هم در مهار پس سیناپسی نقش دارد. تحلیل نورونهای زاینده گابا گابرژیک) در مغز باعث بیماری کره یا داء الرقص میشود که در آن حرکات غیر ارادی پردامنه ای در فرد بیمار ظاهر می شود. یک گروه از مواد دارویی آرام بخش و ضد اضطSراب و خواب آور به نام بنزودی از بین مانند دیازپام عمل گابا را در مغز تشدید و تسهیل می کنند. همین مواد عضلات اسکلتی را نیز منبسط می کنند. بعضی مواد دارویی ضد تشنج و بیهوش کننده به نام باربیتورات مانند فنوباربیتال اثر مهار کننده گابا را تقویت می کند. تاکنون سه نوع گیرنده گابا (A و B و C شناسایی شده است که به ترتیب با افزایش ورود کلر به نورونها، افزایش خروج پتاسیم از آنها و کاهش کلسیم به آنها باعث هیپرپولاریزاسیون غشای نورونها و ایجاد پتانسیل پس سیناپسی مهار کننده میشوند.
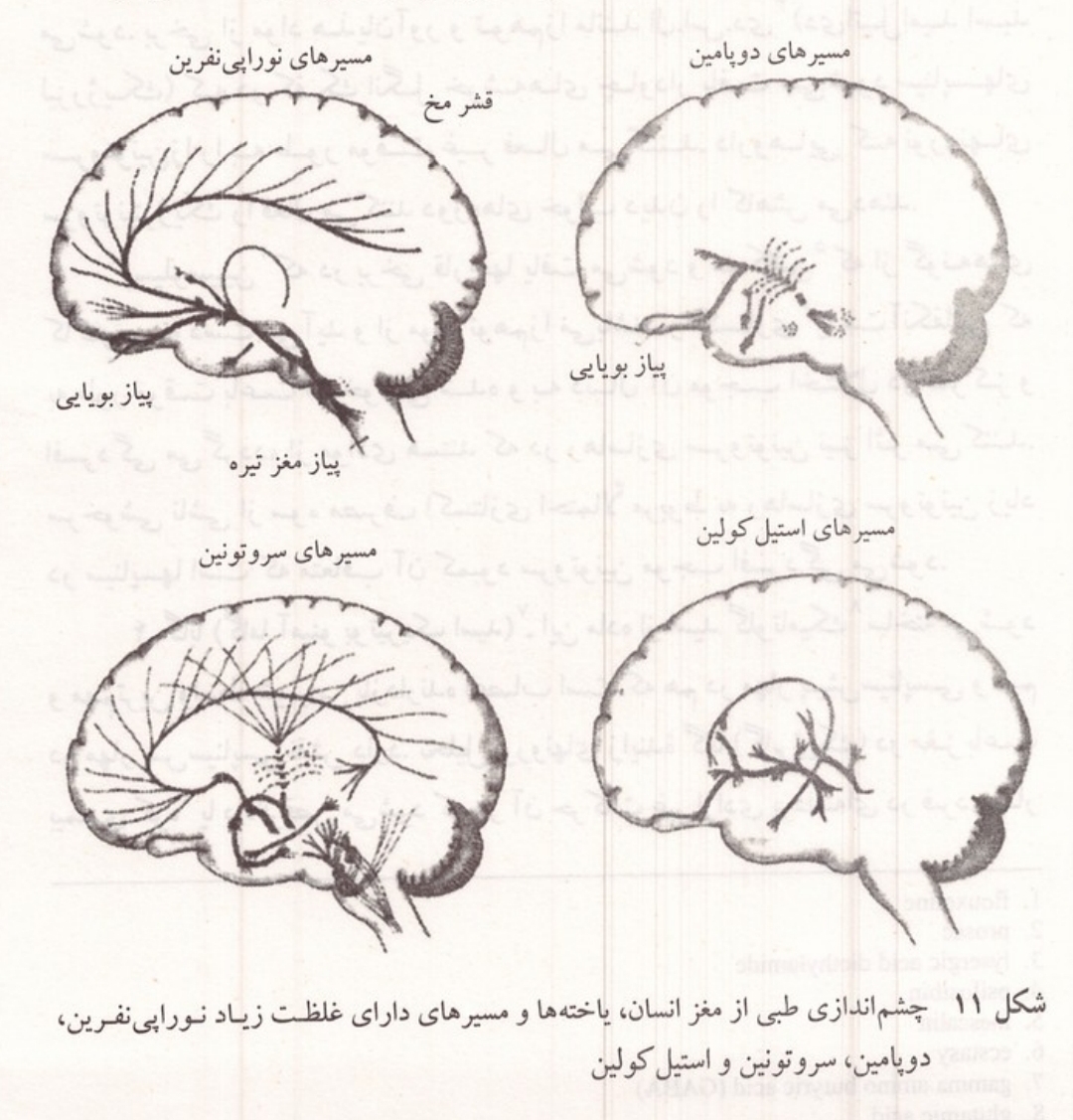
شکل ۱۱: چشم اندازی طبی از مغز انسان؛ یاخته ها و مسیرهای دارای غلظت زیاد نوراپی نفرین، دوپامین، سروتونین و استیل کولین.
د. گلوتامات، آسپارتات، گلیسین
گلوتامات مانند گابا از اسید گلوتامیک به وجود می آید؛ ولی برخلاف گابا میانجی عصبی تحریک کننده است. گلوتامات اصلی ترین میانجی تحریک کننده در سیناپسهای مرکزی است و در حدود سه چهارم كل نورونهای تحریک کننده گلوتاماترژیک هستند. گلوتامات و گابا در سیناپسهای جانوران بسیار ساده نیز شناسایی شده اند. گلوتامات و اسید گلوتامیک در مغز انسان باعث افزایش برانگیختگی و بالا بردن سطح هوشیاری میشوند و به نظر می رسد که در فرایند یادگیری مؤثرند. گلوتامات گابا آسپارتات و گلیسین از گروه اسیدهای آمینه امینواسید) هستند. گلوتامات و آسپارتات از گوارش مواد پروتئینی در روده به وجود می آیند؛ ولی پس از جذب نمیتوانند از سد بین خون و مغز عبور کرده وارد مغز شوند. در عوض مغز خود این مواد را میسازد. برخی از تجربیات جدید نشان داده اند که گلوتامات علاوه بر تحریک نورونها میتواند سلولهای نوروگلیا را نیز دپولاریزه کند. گلوتامات واسطه شیمیایی برخی از نورونهای هیپوتالاموس است که باعث تنظیم ترشحات غده هیپوفیز میشوند. میزان گلوتامات در مایع خارج سلولی مغز بسیار کم است ولی در مواردی مانند ضربه های مغزی و کمبود اکسیژن و کمبود گلوکز که روند برداشت گلوتامات به وسیله نورونها و نورو گلیاها را مختل می سازد افزایش مییابد. افزایش گلوتامات و بعضی از آگونیستهای آن در محیط خارجی نورونها خاصیت سمی دارد و با افزایش ورود کلسیم به نورونها موجب مرگ آنها میشود. در پژوهشهای علوم اعصاب از این سموم تحریک کننده برای تخریب اجسام سلولی نورونها در یک ناحیه بدون آسیب رساندن به مناطق دیگر استفاده میشود. آسپارتات مانند گلوتامات تحریک کننده است. اسید آمینه گلیسین در بسیاری از سیناپسهای نخاع و بخش پایین مغز یک واسطه شیمیایی بازدارنده مهم محسوب میشود و با افزایش ورود کلر باعث مهار نورونهای حرکتی می گردد، به طوری که اختلال در ساخته شدن این ماده باعث انقباض مداوم عضلات می شود.
۶) نوروپپتیدها یا میانجیهای درشت مولکول
یکی از پیشرفتهای مهمی که در نیمه دوم قرن گذشته در بیوشیمی مغز صورت گرفته است کشف نوروپتیدها و گیرنده های آنها از جمله مواد شبه مرفینی و گیرنده های آنها در مغز است. این مواد در اجسام سلولی نورونها ساخته شده به آرامی در آکسونها جابه جا می شوند و سرانجام مانند سایر انتقال دهنده ها از پایانه های سیناپس بیرون می روند. بعضی از این مواد در سایر بافتهای بدن نیز ساخته میشوند؛ ولی سد بین خون و مغز مانع ورود آنها به مغز میشود و نوروپپتیدهایی که در دستگاه عصبی وجود دارند به وسیله یاخته های عصبی مغز و نخاع ساخته میشوند. تعدادی از این مواد پپتیدهای شبه افیونی (اپیوئید) نامیده میشوند که از ماده ای به نام بتالیپوتروپین به وجود می آیند و سه دسته از مواد را به نامهای آندورفینها آنکفالینها و دينورفينها تشكيل می دهند. این مواد با اثر در گیرنده های خود باعث تخفیف درد و ایجاد آرامش عصبی میشوند. قدرت اثر ضد درد مواد شبه مرفینی مغز از مرفین بیشتر است. اثر دارونماها و ضد درد طب سوزنی را به افزایش مواد شبه مرفینی مغز نسبت می دهند. ورزش شدید درد و عوامل استرس زا باعث افزایش ساخته شدن مواد شبه مرفینی در مغز و همچنین افزایش ترشح هورمونهای محرک بخش قشری غده فوق کلیه و كورتيزول میشود. مرفین که از تریاک گرفته میشود از نظر ترکیب شیمیایی با مواد شبه مرفینی مغز متفاوت است؛ ولی بخشی از مولکول آن که در گیرنده های غشای نورونها اثر میکند شبیه مواد شبه مرفینی مغز است. ماده ای به نام نالوکسان باعث جدا شدن مرفین و مواد شبه مرفینی از گیرنده های آنها می شود.
نورو پپتیدهای دیگری در مغز ساخته میشوند که تعدیل کننده و تنظیم کننده حالات روانی بوده در رفتارهای مربوط به گرسنگی سیری تشنگی حالات هیجانی، افسردگی، تغییرات خلقی، حافظه و یادگیری شرکت دارند. اغلب این مواد در سایر نواحی بدن یافت می شوند؛ ولی نقش آنها در مغز با نقشی که در سایر بافتهای بدن دارند متفاوت است. تاکنون دهها نورو پپتید شناسایی شده است. بعضی از آنها علاوه بر مغز در غدد درون ریز نیز ساخته میشوند. کوله سیستو کینین یکی از هورمونهای لوله گوارش است که باعث انقباض کیسه صفرا و کمک به هضم چربیها می شود؛ ولی همین ماده در مغز موجب ایجاد احساس سیری و کاهش اشتها میگردد. هورمون دیگری به نام آنتی دیورتیک که در هیپوتالاموس ساخته میشود و در کلیه ها اثر کرده حجم ادرار ادرار را کاهش میدهد در روند حافظه و یادگیری در مغز نیز شرکت دارد. آنژیوتانسین هورمون دیگری است که با منقبض کردن عضلات صاف دیواره رگها باعث تنگ شدن آنها میشود. همین ماده در مغز عامل ایجاد احساس تشنگی است.
از دیگر نورو پپتیدهایی که در مغز و نخاع ساخته می شوند و بسیاری از آنها در سایر بافتهای بدن نیز وجود دارند میتوان ماده P و نوروپپتید ۷ را ذکر کرد. ماده از جمله در پایانه های اولین نورونهای راه درد در شاخ خلفی نخاع به میزان زیاد وجود دارد و میانجی انتقال درد مزمن است. نوروپپتید ۷ نیز به فراوانی در دستگاه عصبی ساخته میشود و یکی از اعمال آن افزایش اشتها و میل به دریافت غذا است به طوری که آنتاگونیستهای آن ممکن است برای درمان چاقی مؤثر باشند. نورو پپتیدهای دیگری که کانابینوئید نامیده میشوند و آناندامید نیز یکی از آنهاست در بعضی نواحی مغز مانند کورتکس مخچه، بادامه و هیپوکامپ ساخته می شوند. تتراهیدروکانابینول یا THC که ماده مؤثره موجود در حشیش است به گیرنده های آناندامید متصل میشوند و حالتی خلسه مانند ایجاد می کنند.
مواد مؤثر در سیناپسها و چگونگی اثر داروهای روانی
مواد شیمیایی زیادی در گیرنده های غشای سیناپسها اثر میکنند و باعث تحریک یا مهار آنها میشوند. برخی مواد دیگر از آزاد شدن واسطه شیمیایی جلوگیری می کنند یا برعکس مانع از بین رفتن آن در شکاف سیناپس میشوند. این گونه مواد نتایج روانی و رفتاری متفاوتی به بار می آورند و در پژوهشهای بنیادی مربوط به شیمی رفتاری و در روان پزشکی کاربرد دارند. از این مواد برای بررسی نقش انتقال دهنده های عصبی و گیرنده های آنها در اختلالات روانی و رفتاری استفاده می شود. موادی که عمل انتقال دهنده عصبی را تسهیل میکنند همکنش (آگونیست آنها و موادی که باعث بازداری آثار آن میشوند همستیز آنتاگونیست آنها خوانده میشوند. برخی از مواد دارویی ساخته شدن انتقال دهنده عصبی را افزایش می دهند؛ مثلاً ماده ای به نام ال - دوپا دی هیدرو کسی فنیل آلانین که از سد بین خون و مغز عبور می کند؛ در مغز به دوپامین تبدیل شده کمبود آن را جبر میسازد؛ بنابراین ال - دوپا یکی از آگونیستهای دوپامین است. مواد دیگری بر عکس مانع ساخته شدن یک انتقال دهنده می شوند؛ مثلاً پارا کلرو فنیل آلانین مانع ساخته شدن سروتونین می شود و یکی از آنتاگونیستهای آن است. زهر برخی از عنکبوتها باعث آزاد شدن استیل کولین از سیناپسهای کولینرژیک میشود (آگونیست) و توکسین بوتولیک مانع ترشح استیل کولین میشود آنتاگونیست. برخی از مواد شیمیایی و دارویی با اتصال به گیرنده های پس سیناپسی و اشغال آنها مانع اثر کردن انتقال دهنده در آنها می شوند. به این گونه مواد بازدارنده های گیرنده گفته میشود. داروسازان از مجموعه مواد مؤثر در سیناپس برای ساختن داروهای جدید مؤثر در اعصاب و روان استفاده می کنند.
اغلب داروهای ضد روان پریشی بر یک نوع گیرنده دوپامین قرار گرفته آنها را اشغال می کنند و از کار میاندازند. هالوپریدول که دارویی مشهور ضد روان پریشی است به همین طریق عمل میکند. آمفتامینها و کوکائین که اعتیاد آورند از جمله مواد مؤثر در نورونهای دوپامینرژیک هستند ،آمفتامین آزاد شدن دوپامین را افزایش می دهد و کوکائین مانع جذب مجدد آن میشود که در نتیجه با مصرف هر یک از این دو نوع مواد بر مقدار دوپامین در سیناپسها افزوده میشود. مصرف مکرر این مواد در افراد طبیعی باعث بروز نشانه های روان پریشی مانند اسکیزوفرنی می گردد.
بسیاری از داروهای ضد افسردگی از طریق افزایش مقدار نوراپی نفرین عمل میکنند و اغلب مواد دارویی اضطراب زدا گیرنده های گابا را فعال می سازند. فرضیه هایی که در مورد علل بیوشیمیایی برخی از بیماریهای روانی مانند اسکیزوفرنی و افسردگی ارائه شده اند بر اساس این گونه پژوهشهای مربوط به شیمی مغز بنا شده اند. در آن دسته از بیماران مبتلا به اسکیزوفرنی که با مصرف داروهای ضد روان پریشی بهبود می یابند از غلظت هو مو وانیلیک اسید (HVA) در خون آنان که نتیجه تجزیه دوپامین است کاسته میشود. بالا بودن فعالیت نورونهای دوپامینرژیک و نورآدرنرژیک در شیدایی و بیش فعالی و پایین بودن فعالیت آنها در افسردگی از جمله شواهدی است که در تأیید این فرضیه ها ذکر شده است. داروهای ضد افسردگی سه حلقه ای و داروهای بازدارنده مونو آمین اکسید از که در برخی بیماران افسرده اثر بهبود بخش دارند تا حدی مؤید نقش کاتکولامینها و سروتونین در این بیماریهاست. با وجود آنچه گفته شد باید به پیچیدگی کار مغز و تداخل اثر انتقال دهنده ها و گیرنده های آنها و روابط پیچیده ای که بین انتقال دهنده های مختلف با یکدیگر و با گیرنده ها وجود دارد توجه داشت و به این قبیل نظریه های بیوشیمیایی که کار مغز را بیش از حد ساده نشان میدهند با دید انتقادی نگاه کرد. در حال حاضر اعتقاد بر این است که مهمترین موادی که در علوم اعصاب و دارو درمانی بیماریهای روانی مورد استفاده قرار میگیرند به شرح زیر عمل می کنند:
- مواد ضد درد: این مواد به دو گروه اصلی دردزداهای افیونی و دردزداهای غیر افیونی تقسیم میشوند. ترکیبات افیونی مانند مرفین کدئین، هروئین و متادون بر گیرنده های ویژه ای که قبلاً در مبحث مواد شبه افیونی مغز شرح داده ایم قرار می گیرند. دردزداهای غیر افیونی از طریق کاهش تولید پروستاگلاندینها که از عوامل اصلی ایجاد درد در بافتها هستند عمل می کنند.
- مواد ضد افسردگی: این نظر که افسردگی ممکن است نتیجه کاهش فعالیت نورونهای ترشح کننده سروتونین و نوراپی نفرین باشد ناشی از این است که داروهایی مانند رزرپین که مانع ترشح آنها میشوند اغلب افسردگی ایجاد می کنند و بسیاری از بیماران افسرده با استفاده از داروهایی که فعالیت نورونهای نورآدرنرژیک و سروتونرژیک را افزایش میدهند بهبود می یابند. مهار کننده های مونو آمین اکسیداز که مانع تخریب این انتقال دهنده ها میشود و ضد افسردگیهای سه حلقه ای مانند ایمی پرامین و آمی تریپتیلین که مانع باز جذب آنها میشود عمده ترین این داروها است. بعضی بیماران به طور متناوب دچار حالات افسردگی و شیدایی (مانیا) می شوند که آن را اختلال دو قطبی مینامند. کربنات لیتیوم که یک ماده طبیعی معدنی است در افراد مبتلا به افسردگی دو قطبی کاربرد دارد. لیتیوم جانشین سدیم شده مانع تحریک شدید و بروز دوره های شیدایی (مانیا) در افراد بیمار می شود.
- مواد ضد اسکیزوفرنی: این مواد گیرنده های دوپامین را مهار کرده مانع اثر دوپامین در آنها میشوند و آنتاگونیست دوپامین هستند. در اواسط قرن بیستم دارویی به نام کلرپرومازین ساخته شد و برای آرام کردن بیماران آشفته و برانگیخته مبتلا به اسکیزوفرنی مورد استفاده قرار گرفت. این ماده که به عنوان آرام بخش نیز کاربرد دارد، همراه با سایر آنتاگونیستهای دوپامین در بیماران اسکیزوفرن مصرف می شود.
- مواد بالابرنده سطح هوشیاری: برخی مواد مانند نیکوتین، آمفتامین و کوکائین باعث برانگیختگی و افزایش سطح هوشیاری و بالا بردن دقت و توجه می شوند. اثر نيكوتين مربوط به تحریک یک نوع از گیرنده های کولینرژیک گیرنده های نیکوتینی) است؛ ولی این ماده همراه با سایر مواد موجود در دخانیات دارای آثار جانبی نامطلوب زیادی است. اثر آمفتامین و کوکائین را در افزایش کاتکولامینها در سیناپسها قبلاً ذکر کرده ایم. افزایش فعالیت نورونهای نورآدرنرژیک باعث تشدید فعالیت و افزایش هوشیاری میشود و دارای آثار رفتاری بسیار متعدد و متنوعی است. مصرف دراز مدت مواد بالا برنده سطح هوشیاری عوارضی شبیه اسکیزوفرنی را ایجاد می کند.
- مواد توهم زا: اغلب این مواد آنتاگونیستهای سروتونین هستند. مشتقات حشیش که در گیاه شاهدانه هندی وجود دارد و ال اس دی و پسیلوسیبین که در بعضی از کفکها و قارچها یافت میشود از این دسته مواد هستند. این مواد ممکن است علاوه بر توهم باعث ایجاد اضطراب و ترس شوند. مصرف این مواد سروتونین مغز را کاهش میدهد و در نتیجه مدارهای بازدارنده رؤیا در مغز از کار افتاده، توهمات رؤیا گونه ایجاد میشوند. تعداد دیگری از مواد توهم زا مانند مسکالین که در یک نوع گیاه کاکتوس یافت میشود همکنش آگونیست) نورآدرنالین هستند و بعضی دیگر در فعالیت سایر انتقال دهنده های عصبی و گیرنده های آنها اثر می کنند.
- مواد بی حس کننده و بیهوش کننده: بی حس کننده های موضعی ایجاد و هدایت موج عصبی را متوقف میکنند. عمل این مواد با کاهش نفوذ پذیری غشا نسبت به سدیم صورت میگیرد و برگشت پذیر است. بسته شدن مجاری سدیم در غشای عصبی، ایجاد موج عصبی و هدایت آن را در طول عصب غیر ممکن می سازد. بیهوش کننده های عمومی که به دو صورت استنشاقی و تزریقی به کار میروند در سطح سیناپسهای مرکزی مغز و نخاع عمل میکنند و سطح هوشیاری را پایین می آورند. اثر اختصاصی این مواد از کار انداختن سیناپسهای تحریک کننده تشکیلات تورینه ای تنه مغز است که در نتیجه آن فعالیت نورونهای قشر مخ کاهش یافته سطح هوشیاری پایین می آید. مراحل بیهوشی با کاهش آگاهی از درد آغاز میشود و به تدریج به از بین رفتن هوشیاری و تغییر انعکاسها و اعمال نباتی بدن منجر میشود. در بیهوشی عمیق فعالیت بصل النخاع تضعیف شده، احتمال اختلالات تنفسی و قلبی - عروقی افزایش می یابد. مواد بی حس کننده و بیهوش کننده با قابلیت انحلال خود در چربیها به سرعت از غشای عصبی عبور کرده اثر خود را نمایان می سازند.
- مواد ضد اضطراب: بنزودیازپینها که مهمترین داروهای ضد اضطراب هستند و دیازپام (والیوم) نمونه ای از آنهاست در یکی از انواع گیرنده های گابا اثر می کنند و با توجه به اینکه گابا از واسطه های مهار کننده است موجب کاهش اضطراب می شوند. چنان که قبلاً نیز اشاره شده است باربیتوریکها یک گروه از اضطراب زداها هستند که آنها نیز از طریق افزایش فعالیت گابا اثر می کنند.
برخی از سایر روان گردانها
همان طور که قبلاً ذکر شد اثر بعضی از مواد دارویی و اعتیاد آور بر مغز شبیه اثر فراورده های نورونهاست. این مواد فعالیت نوروترانسمیترها را تشدید میکنند و یا از فعالیت آنها می کاهند. تغییرات زیان آوری که اغلب این مواد در مغز ایجاد میکنند حتی پس از قطع مصرف آنها ادامه می یابد. عمده ترین این مواد به شرح زیر است:
- نیکوتین: این ماده آلکالوئید موجود در دود سیگار است که به سرعت وارد مغز شده بر گیرنده های نیکوتینی استیل کولین که در بسیاری از نواحی مغز مانند قشر مخ مخچه و بخشهایی از مغز که در کنترل حرکات و تفکر و هیجانها اثر می کند یافت می شوند.
- حشیش: قبلاً گفته شده است که تتراهیدروکانابینول بر گیرنده های کانابینوئیدها اثر کرده و توجه و هماهنگی اندامها را کاهش میدهد و در دراز مدت بی قیدی احساس ملال اختلال حافظه و کاهش قدرت تمرکز فکری به بار می آورد. بررسی در جانوران نشان داده است که سوء مصرف کانابیس باعث زوال نورونها و کاهش ارتباط آنها در هیپوکامپ و در نتیجه اختلال در یادگیری و حافظه می شود.
- کوکائین: کوکائین از برگ گیاه کوکا که در آمریکای جنوبی می روید به دست می آید و یک محرک قوی است. پودر سفیدرنگ کوکائین به صورتهای مختلف استنشاقی، تزریقی و یا به صورت سیگار مصرف می شود ، اشتها را کنم می کند و به ویژه در نقاطی از مغز که دارای نورونهای دوپامینرژیک است، اثر کرده و باعث تغییرات خلقی میشود. در این حالت خون بخشهایی از مغز که اعمال برتر عصبی مانند تفکر و حافظه را به عهده دارند کاهش می یابد. بنابر برخی تحقیقات، کو کائین تعداد گیرنده های دوپامین در مغز را کاهش می دهد و این تغییر عامل احساس ناراحتی شدید در هنگام ترک اعتیاد به کوکائین است. استفاده مکرر از کوکائین توانایی مغز را در مصرف گلوکز که سوخت اصلی مغز است کاهش می دهد.
- هروئین: هروئین مانند سایر مشتقات تریاک (اپیاتها) با اثر بر گیرنده های اوپیوئیدی مغز که بیشتر در کف بطن چهارم و اطراف قنات سيلويوس و بصل النخاع وجود دارند، باعث دردزدایی و سرخوشی میشود. همین طور مردمک چشم را تنگ کرده ممکن است اختلالات تنفسی و تهوع ایجاد کند و خطر سکته مغزی و ایجاد لخته در خون را افزایش میدهد. حتی یک بار مصرف هروئین اعتیاد آور است.
- متادون: یک ماده جانشین برای ترک اعتیاد به هروئین است که حالت سرخوشی ناشی از مصرف هروئین را تولید نمیکند ولی عوارض ترک آن را تحمل پذیرتر می کند.
- آمفتامینها و متامفتامینها: این مواد باعث رهاسازی دوپامین و در نتیجه احساس شدید قدرت و لذت میشود و برای بیدار ماندن طولانی و افزایش توان کار و کاهش اشتها به کار رفته اند. آمفتامینها ضربان قلب و حرکات تنفس و فشار خون و قطر مردمک را افزایش میدهند و در پزشکی جز در موارد نادر مانند درمان نارکولپسی (حمله خواب کاربرد ندارند. از عوارض جانبی مصرف این مواد می توان اضطراب، اختلالات خواب بی اشتهایی گیجی و منگی و توهم و هذیان و اختلالات رفتاری را نام برد.
رسانههای تکمیلی فصل
پادکست
ویدیو
آزمونک فصل چهارم (سوالات تشریحی)
دانش خود را بر اساس مطالب این فصل بسنجید.
۱. سیناپس چیست و رایجترین انواع آن از نظر محل اتصال کدامند؟
نمایش پاسخ
پاسخ: سیناپس، محل ارتباط یک نورون با نورون دیگر است. بیشترین تعداد سیناپسها از نوع **آکسون به دندریت** و یا **آکسون به جسم سلولی** است.
۲. تفاوت اصلی سیناپس الکتریکی و شیمیایی از نظر ساختار (فاصله غشاها) و جهت انتقال پیام را توضیح دهید.
نمایش پاسخ
پاسخ: در سیناپس **الکتریکی**، غشای دو نورون به یکدیگر میچسبد و فاصلهای وجود ندارد؛ انتقال بدون واسطه و اغلب در **هر دو جهت** امکانپذیر است.
در سیناپس **شیمیایی**، بین دو غشا فاصلهای به نام **شکاف سیناپسی** (۲۰۰ تا ۳۰۰ آنگستروم) وجود دارد؛ انتقال با واسطه شیمیایی و **یکطرفه** است.
۳. چرا سیناپسهای شیمیایی نسبت به سیناپسهای الکتریکی یک مزیت عملکردی مهم دارند؟
نمایش پاسخ
پاسخ: مزیت اصلی، امکان **بازداری و مهار** نورون پسسیناپسی است. این قابلیت در سیناپسهای الکتریکی که همواره تحریککننده هستند، وجود ندارد.
۴. نقش یون کلسیم (Ca2+) در فرآیند انتقال سیناپسی چیست؟
نمایش پاسخ
پاسخ: هنگامی که موج عصبی (دپولاریزاسیون) به دکمه سیناپسی میرسد، باعث میشود یونهای **کلسیم به پایانه سیناپس وارد شوند**. ورود کلسیم باعث اتصال غشای پیشسیناپسی به کیسههای حاوی انتقالدهنده شده و موجب آزاد شدن محتویات این کیسهها به شکاف سیناپس میگردد.
۵. پدیده «خستگی سیناپس» (Synaptic Fatigue) را توضیح دهید و بگویید چرا تار عصبی خسته نمیشود اما سیناپس خسته میشود؟
نمایش پاسخ
پاسخ: تحریک مکرر و شدید نورونها میتواند باعث خستگی سیناپس شود. در این حالت، نورون فرصت **بازسازی انتقالدهندههای عصبی** را پیدا نمیکند و سیناپس بر اثر **کمبود این مواد** از کار میافتد. در حالی که تار عصبی (به شرط رعایت زمان تحریکناپذیری) دچار خستگی نمیشود.
۶. مکانیسم یونی سیناپسهای **تحریک کننده** (که منجر به EPSP میشود) چیست؟
نمایش پاسخ
پاسخ: در این سیناپسها، انتقالدهنده به گیرندههای غشای پسسیناپسی متصل شده و باعث نفوذپذیر شدن این غشا نسبت به **یونهای سدیم (Na+)** میشود. ورود یونهای سدیم باعث کاهش پتانسیل غشا (دپولاریزاسیون) و تحریک آن میگردد.
۷. مکانیسم یونی سیناپسهای **بازدارنده** (که منجر به IPSP میشود) چیست؟ (دو مکانیسم ممکن را ذکر کنید)
نمایش پاسخ
پاسخ: انتقالدهنده در این سیناپسها نفوذپذیری غشا را نسبت به سدیم تغییر نمیدهد، بلکه آن را نسبت به **یونهای پتاسیم (K+)** و یا **یونهای کلر (Cl-)** افزایش میدهد.
۱. افزایش نفوذ به پتاسیم باعث **خروج** پتاسیم از سلول میشود.
۲. افزایش نفوذ به کلر باعث **ورود** کلر به سلول میشود.
هر دو حالت منجر به افزایش پتانسیل آرامش (هیپرپولاریزاسیون) و مهار نورون میشوند.
۸. «مهار پیش سیناپسی» (Presynaptic Inhibition) را توضیح دهید و بگویید چگونه مانع انتقال پیام میشود؟
نمایش پاسخ
پاسخ: این پدیده به واسطه سیناپسهای **آکسون به آکسون** صورت میگیرد. یک نورون رابط، انتقالدهندهای آزاد میکند که نفوذپذیری غشای نورون تحریککننده (پیشسیناپسی) را نسبت به **یون کلسیم کاهش میدهد**. در نتیجه، با رسیدن موج عصبی، کلسیم کافی وارد پایانه نشده، خروج انتقالدهنده کاهش مییابد و موج عصبی متوقف میشود.
۹. نقش «گیرندههای پیش سیناپسی» (Autoreceptors) چیست؟
نمایش پاسخ
پاسخ: این گیرندهها نسبت به انتقالدهندهای که از **همان پایانه عصبی** آزاد میشود، حساسیت دارند. آنها با یک **چرخه بازخورد منفی** باعث تنظیم مقدار انتقالدهنده شده و مانع آزاد شدن بیش از حد آن میشوند.
۱۰. تفاوت گیرندههای «یونوتروپیک» (Ionotropic) و «متابوتروپیک» (Metabotropic) را از نظر ساختار، سرعت و مکانیسم عمل شرح دهید.
نمایش پاسخ
یونوتروپیک: این گیرندهها خودشان **کانال یونی** (وابسته به لیگاند) هستند. اثر نوروترانسمیتر مستقیماً باعث تغییر شکل و باز شدن مجرا برای عبور یون میشود. این فرآیند **سریع** است.
متابوتروپیک: این گیرندهها مستقیماً کانال را باز نمیکنند، بلکه اثری **کندتر ولی طولانیتر** دارند. اثر نوروترانسمیتر یک سری **تغییرات شیمیایی** (مانند فعال کردن آدنوزین مونوفسفات حلقوی و پروتئین کیناز) را راهاندازی میکند که در نهایت به تغییر پتانسیل غشا منجر میشود.
۱۱. EPSP و IPSP مخفف چیست و هر کدام چه تأثیری بر پتانسیل آرامش غشای پسسیناپسی دارند؟
نمایش پاسخ
EPSP (پتانسیل پسسیناپسی تحریککننده):** پایانه تحریککننده پتانسیل آرامش غشای پسسیناپسی را **کاهش** میدهد (آن را به آستانه تحریک نزدیکتر میکند).
IPSP (پتانسیل سیناپسی بازدارنده):** پایانه بازدارنده پتانسیل آرامش غشای پسسیناپسی را **افزایش** میدهد (آن را از آستانه تحریک دورتر میکند).
۱۲. تفاوت «انتقال دهنده عصبی» و «تعدیل کننده عصبی» از نظر دامنه اثر چیست؟
نمایش پاسخ
پاسخ: **انتقالدهنده** در سطح یک سیناپس اثر میکند و **منطقه عمل محدودتری** دارد. **تعدیلکنندههای عصبی** در مغز **میدان عمل وسیعتری** دارند و میزان ترشح آنها بیشتر است.
۱۳. استیل کولین در کدام سه نقطه اصلی در بدن به عنوان انتقالدهنده عمل میکند؟
نمایش پاسخ
پاسخ: ۱. انتقالدهنده منحصر به فرد همه سیناپسهای **عصب به عضلات اسکلتی**.
۲. در **عقدههای خودکار** (اتونوم).
۳. در پایانههای سیناپسی اعصاب **پاراسمپاتیک** (و بخشی از سمپاتیک).
۱۴. چرا استیل کولین بر عضلات اسکلتی اثر تحریکی، اما بر عضلات قلب اثر بازدارندگی دارد؟
نمایش پاسخ
پاسخ: این تفاوت به **نوع گیرندههای پسسیناپسی** مربوط است.
- در **عضلات اسکلتی**، در مجاری **سدیم** اثر کرده و باعث دپولاریزاسیون (تحریک) میشود.
- در **قلب**، گیرندهها مربوط به مجاری **پتاسیم** بوده و اثر استیل کولین باعث افزایش پتانسیل آرامش (بازدارندگی) میشود.
۱۵. دو نوع اصلی گیرندههای استیل کولین چه نام دارند؟ کدام یک یونوتروپیک و کدام یک متابوتروپیک است؟
نمایش پاسخ
پاسخ: ۱. **گیرندههای نیکوتینی** (فعالشونده با نیکوتین).
۲. **گیرندههای موسکارینی** (فعالشونده با موسکارین).
گیرندههای **نیکوتینی** از نوع **یونوتروپیک** (همیشه تحریکی) هستند.
گیرندههای **موسکارینی** از نوع **متابوتروپیک** (تحریکی یا مهاری) هستند.
۱۶. بیماری آلزایمر با اختلال در کدام سیستم انتقالدهنده در مغز مرتبط است؟
نمایش پاسخ
پاسخ: در بیماری آلزایمر، نورونهای **استیل کولینرژیک** مغز دچار کمکاری و مرگ تدریجی میشوند.
۱۷. استیل کولین پس از اثر چگونه غیرفعال میشود؟ و آفتکشها (ترکیبات ارگانوفسفات) چگونه این روند را مختل میکنند؟
نمایش پاسخ
پاسخ: استیل کولین توسط آنزیم **استیل کولین استراز** تجزیه و غیرفعال میشود.
آفتکشها و گازهای عصبی با **خنثی کردن اثر این آنزیم**، باعث طولانی شدن فعالیت سیناپسهای کولینرژیک شده که به تشنج و اغما منجر میگردد.
۱۸. کاتکولامینها شامل چه موادی هستند و از کدام اسید آمینه ساخته میشوند؟
نمایش پاسخ
پاسخ: شامل **دوپامین**، **نوراپی نفرین** (نورآدرنالین) و **اپی نفرین** (آدرنالین) هستند. همگی از اسید آمینه **تیروزین** ساخته میشوند.
۱۹. بیماری پارکینسون چگونه در اثر اختلال در سیستم دوپامینرژیک ایجاد میشود؟ (به مسیر نیگرواستریاتال اشاره کنید)
نمایش پاسخ
پاسخ: بر اثر آسیب و تحلیل تدریجی نورونهای دوپامینرژیک **هسته سیاه** (در مغز میانی). این امر باعث کاهش دوپامین در **اجسام مخطط** (هستههای قاعدهای مغز) میشود (که به آن مسیر نیگرواستریاتال میگویند). این کمبود دوپامین سبب پرکاری نورونهای استیل کولینرژیک در آن ناحیه شده و لرزش و سختی عضلات ایجاد میشود.
۲۰. نقش دوپامین در بیماری اسکیزوفرنی (روانگسیختگی) چیست؟ و داروهای نورولپتیک چگونه آن را درمان میکنند؟
نمایش پاسخ
پاسخ: به نظر میرسد **فعالیت بیش از حد** نورونهای دوپامینرژیک (در مسیر مزوکورتیکال) و یا حساسیت زیاد گیرندههای آنها، عامل بخشی از نشانههای اسکیزوفرنی باشد.
داروهای نورولپتیک (مانند هالوپریدول) **گیرندههای دوپامین را بازداری (مهار)** کرده و موجب کاهش فعالیت این نورونها و تخفیف علائم میشوند.
۲۱. کاتکولامینها (و دوپامین) توسط کدام دو آنزیم اصلی تجزیه و غیرفعال میشوند؟
نمایش پاسخ
پاسخ: ۱. **مونو آمین اکسیداز (MAO)** (آنزیم داخل نورون).
۲. **کاتکول اومتیل ترانسفراز (COMT)** (آنزیم خارج نورون).
۲۲. نورونهای نورآدرنرژیک (تولیدکننده نوراپی نفرین) در کدام بخش تنه مغزی متمرکز شدهاند و نقش اصلی آنها چیست؟
نمایش پاسخ
پاسخ: بیشتر در تنه مغزی، از جمله **هسته نیلی (لوکوس سرولئوس)** در پل مغز قرار دارند. نقش اصلی آنها در **بیداری و افزایش سطح برانگیختگی** و بالا بردن فعالیت نورونهای مغز است.
۲۳. تفاوت عملکردی گیرندههای آلفا و بتای نوراپی نفرین در چیست؟
نمایش پاسخ
پاسخ: در اغلب موارد، اثر نوراپی نفرین در گیرندههای **آلفا از نوع تحریکی** و در گیرندههای **بتا از نوع مهاری** است. (قلب و روده استثنا هستند).
۲۴. سروتونین از کدام اسید آمینه ساخته میشود و در کدام بخش مغز به فراوانی یافت میشود؟
نمایش پاسخ
پاسخ: از اسید آمینه **تریپتوفان** ساخته میشود. در **هسته سجافی (رافه)** در تنه مغزی به فراوانی ساخته میشود.
۲۵. سروتونین در چه اعمالی نقش دارد؟ (حداقل ۳ مورد)
نمایش پاسخ
پاسخ: سروتونین در برقراری حالت **خواب**، مهار مسیرهای انتقال **درد**، و تنظیم **رفتارهای تغذیه** و **نوسانات خلق (افسردگی)** نقش دارد.
۲۶. داروهای ضد افسردگی مانند «فلوئوکستین (پروزاک)» و «بازدارندههای MAO» چگونه باعث بهبود افسردگی میشوند؟
نمایش پاسخ
پاسخ: **بازدارندههای MAO** مانع تجزیه سروتونین (و کاتکولامینها) میشوند.
**فلوئوکستین (پروزاک)** به صورت اختصاصی مانع **بازجذب** سروتونین میشود.
هر دو دارو در نهایت باعث افزایش مقدار سروتونین در سیناپسها میشوند.
۲۷. مواد توهمزا مانند LSD و اکستازی چگونه بر سیستم سروتونین اثر میگذارند؟
نمایش پاسخ
پاسخ: **LSD** سیناپسهای سروتونینزا را به طور موقت **غیر فعال** میکند.
**اکستازی** باعث **رهاسازی سروتونین زیاد** در سیناپسها (ایجاد سرخوشی) و متعاقب آن **کمبود سروتونین** (ایجاد افسردگی) میشود.
۲۸. «گابا» (GABA) چیست، از چه مادهای ساخته میشود و چه نقشی دارد؟
نمایش پاسخ
پاسخ: گابا (گاما آمینو بوتیریک اسید) از **اسید گلوتامیک** ساخته میشود. این ماده، **مهمترین واسطه شیمیایی بازدارنده** اعصاب است که هم در مهار پیشسیناپسی و هم در مهار پسسیناپسی نقش دارد.
۲۹. بیماری «کره» (داءالرقص) ناشی از اختلال در کدام سیستم انتقالدهنده است؟
نمایش پاسخ
پاسخ: ناشی از **تحلیل نورونهای زاینده گابا (گابرژیک)** در مغز است که منجر به حرکات غیر ارادی پردامنه میشود.
۳۰. داروهای آرامبخش مانند «بنزودیازپینها (دیازپام)» و «باربیتوراتها» چگونه عمل میکنند؟
نمایش پاسخ
پاسخ: هر دو گروه دارویی، عمل **گابا** (انتقالدهنده مهاری) را در مغز **تشدید و تسهیل** میکنند و از این طریق اثر آرامبخش، ضد اضطراب و خوابآور دارند.
۳۱. «گلوتامات» چیست، چه نقشی دارد و چرا «اصلیترین» میانجی در نوع خود محسوب میشود؟
نمایش پاسخ
پاسخ: گلوتامات (برخلاف گابا) میانجی عصبی **تحریککننده** است. این ماده **اصلیترین** میانجی تحریککننده در سیناپسهای مرکزی است و حدود سهچهارم کل نورونهای تحریککننده، گلوتاماترژیک هستند.
۳۲. چرا گلوتامات و آسپارتات موجود در غذای هضم شده (پروتئینها) نمیتوانند وارد مغز شوند؟
نمایش پاسخ
پاسخ: زیرا پس از جذب از روده، نمیتوانند از **سد بین خون و مغز** عبور کنند. به همین دلیل مغز خود این مواد را میسازد.
۳۳. افزایش گلوتامات در مایع خارج سلولی (مثلاً در ضربه مغزی یا کمبود اکسیژن) چه خطری برای نورونها دارد؟
نمایش پاسخ
پاسخ: افزایش گلوتامات خاصیت **سمی** دارد و با افزایش ورود **کلسیم** به نورونها، موجب **مرگ نورونها** میشود (Excitotoxicity).
۳۴. نقش اسید آمینه «گلیسین» (Glycine) در نخاع چیست؟
نمایش پاسخ
پاسخ: گلیسین در بسیاری از سیناپسهای نخاع و بخش پایین مغز، یک واسطه شیمیایی **بازدارنده** مهم محسوب میشود. این ماده با افزایش ورود **کلر**، باعث مهار نورونهای حرکتی میگردد.
۳۵. نوروپپتیدها (میانجیهای درشت مولکول) در کجای نورون ساخته میشوند و چگونه به پایانه میرسند؟
نمایش پاسخ
پاسخ: این مواد در **اجسام سلولی نورونها** ساخته شده، به آرامی در **آکسونها جابجا میشوند** و سرانجام از پایانههای سیناپس بیرون میروند.
۳۶. پپتیدهای شبه افیونی (اپیوئید) مغز شامل چه دستههایی هستند و نقش اصلی آنها چیست؟
نمایش پاسخ
پاسخ: شامل سه دسته **آندورفینها**، **آنکفالینها** و **دینورفینها** هستند. نقش اصلی آنها **تخفیف درد** و ایجاد **آرامش عصبی** است.
۳۷. مرفین (گرفته شده از تریاک) چگونه اثر ضد درد خود را اعمال میکند؟
نمایش پاسخ
پاسخ: مرفین از نظر شیمیایی با مواد شبهمرفینی مغز متفاوت است، اما بخشی از مولکول آن شبیه این مواد بوده و بر **گیرندههای غشایی اپیوئیدی** در مغز اثر میکند.
۳۸. نقش نوروپپتید «کوله سیستو کینین» (CCK) در لوله گوارش و در مغز چیست؟
نمایش پاسخ
پاسخ: در **لوله گوارش** (به عنوان هورمون) باعث انقباض کیسه صفرا و کمک به هضم چربیها میشود.
در **مغز** (به عنوان نوروپپتید) موجب ایجاد **احساس سیری** و کاهش اشتها میگردد.
۳۹. نقش نوروپپتید «ماده P» (Substance P) در نخاع چیست؟
نمایش پاسخ
پاسخ: ماده P در پایانههای اولین نورونهای راه درد در شاخ خلفی نخاع به میزان زیاد وجود دارد و میانجی انتقال **درد مزمن** است.
۴۰. «آناندامید» چیست و ماده مؤثره حشیش (THC) چگونه بر آن اثر میگذارد؟
نمایش پاسخ
پاسخ: آناندامید یک نوروپپتید از نوع **کانابینوئید** است که در نواحی مختلف مغز (مانند هیپوکامپ) ساخته میشود. **THC** (تتراهیدروکانابینول) که ماده مؤثره حشیش است، به **گیرندههای آناندامید** متصل شده و حالتی خلسهمانند ایجاد میکند.
۴۱. «آگونیست» (Agonist) و «آنتاگونیست» (Antagonist) را در داروشناسی سیناپسی تعریف کنید.
نمایش پاسخ
آگونیست (همکنش):** موادی که عمل انتقال دهنده عصبی را **تسهیل** میکنند (مانند ال-دوپا برای دوپامین).
آنتاگونیست (همستیز):** موادی که باعث **بازداری** آثار انتقال دهنده عصبی میشوند (مانند پارا کلرو فنیل آلانین برای سروتونین).
۴۲. کوکائین و آمفتامین چگونه بر سیناپسهای دوپامینرژیک اثر میگذارند؟
نمایش پاسخ
پاسخ: **آمفتامین**، **آزاد شدن** دوپامین را افزایش میدهد.
**کوکائین**، مانع **جذب مجدد (بازجذب)** دوپامین میشود.
در نتیجه هر دو ماده، مقدار دوپامین را در سیناپسها به شدت افزایش میدهند.
۴۳. مکانیسم عمل داروهای ضد افسردگی سه حلقهای (مانند ایمیپرامین) و مهارکنندههای مونو آمین اکسیداز (MAO) چیست؟
نمایش پاسخ
پاسخ: هر دو گروه داروهایی هستند که فعالیت نورونهای نورآدرنرژیک و سروتونرژیک را افزایش میدهند.
- **مهارکنندههای MAO:** مانع **تخریب** این انتقالدهندهها توسط آنزیم MAO میشوند.
- **ضد افسردگیهای سهحلقهای:** مانع **بازجذب** این انتقالدهندهها به پایانه پیشسیناپسی میشوند.
۴۴. کربنات لیتیوم چگونه به درمان «اختلال دو قطبی» (افسردگی-شیدایی) کمک میکند؟
نمایش پاسخ
پاسخ: لیتیوم در افراد مبتلا به اختلال دو قطبی کاربرد دارد. این ماده **جانشین سدیم** شده و مانع تحریک شدید و بروز **دورههای شیدایی (مانیا)** در فرد بیمار میشود.
۴۵. مکانیسم عمل مواد بیحس کننده موضعی (مانند پروکائین) و بیهوش کننده عمومی چیست؟
نمایش پاسخ
بیحس کننده موضعی:** با **کاهش نفوذپذیری غشا نسبت به سدیم** (بستن مجاری سدیم)، مانع ایجاد و هدایت موج عصبی در طول عصب میشود.
بیهوش کننده عمومی:** در سطح سیناپسهای مرکزی مغز عمل کرده و با از کار انداختن سیناپسهای تحریک کننده **تشکیلات تورینهای** تنه مغز، فعالیت قشر مخ و سطح هوشیاری را پایین میآورد.
۴۶. مواد ضد اضطراب (مانند دیازپام) و مواد مخدر (مانند هروئین) به ترتیب بر کدام سیستمهای انتقالدهنده اثر میگذارند؟
نمایش پاسخ
مواد ضد اضطراب (بنزودیازپینها):** در یکی از انواع گیرندههای **گابا (GABA)** اثر کرده و چون گابا مهارکننده است، موجب کاهش اضطراب میشوند.
هروئین (اپیاتها):** با اثر بر **گیرندههای اوپیوئیدی (شبه افیونی)** مغز، باعث دردزدایی و سرخوشی میشود.